


Aunque todavía con muchas reservas, la sociedad va prestando creciente atención a esa realidad insoslayable que es fruto de nuestra intrínseca fragilidad biológica: la discapacidad intelectual. No ha sido líder, precisamente, la comunidad neurocientífica en el estudio de esta condición humana. Las primeras aportaciones vinieron del campo de la psicopedagogía y la neuropsicología. Y aquí habríamos de rendir un emocionado homenaje a esos “maestros de escuela” que trataban de desentrañar, y lo conseguían, el mejor modo de enseñar a hablar, o a leer, o a manejarse de manera independiente a niños con discapacidad intelectual, sin esperar a que nadie les dijera el origen del problema.
El reto de la discapacidad intelectual para la neurociencia presenta unas proporciones casi inconmensurables. Primero, por la amplísima diversidad de causas que la provocan —físicas, metabólicas, genéticas, etc. Segundo, por la enorme variedad deformas en que se manifiesta. Tercero, porque en ella se ven implicados los grandes gigantes de la esencia humana: su cognición, su raciocinio, su determinación, sus sentimientos, su conducta. Y en el centro de todo ello, el cerebro.
No debemos ver a la discapacidad intelectual como meramente receptora de lo que la neurociencia le aporta. como neurocientífico y como persona estrechamente vinculada al mundo de la discapacidad intelectual, me gusta contemplarla como realidad que ofrece a la neurociencia nuevas oportunidades para desarrollar y ampliar su conocimiento. El error de un gen, por ejemplo, causante de una determinada forma de discapacidad intelectual, nos permite descubrir mecanismos insospechados en la neuroquímica cerebral o en el funcionamiento normal de la vida de las neuronas, de la glía o de los astrocitos que conforman los normales procesos cerebrales.
Quiero ahora concretar el reto actual de la discapacidad intelectual en el ámbito de la neurociencia mostrando algunos perfiles que considero particularmente incisivos para comprender el mutuo beneficio que ambas entidades se proporcionan. Y lo haré de la mano de aquella que mejor conozco: el síndrome de Down. No es, precisamente, la más sencilla pero sí es, ciertamente —al menos ante la sociedad— la imagen insignia de la discapacidad intelectual. Los perfiles van a utilizar unos elementos clave que utilizaré a modo de ejemplos:
a) la actividad de un gen,
b) el valor de los modelos animales,
c) la modulación neuroquímica,
d) neuroimagen: redes funcionales
La acción de un gen. Modelos animales
Puesto que el síndrome de Down es una trisomía de los genes del 21, está justificado que busquemos en ellos a los responsables, al menos iniciales, de la dismorfia cerebral. Uno de ellos es el gen DYRK1A, ampliamente estudiado por neurocientíficos españoles. Codifica una tirosina cinasa.
Para conocer e ilustrar las consecuencias de una aneuploidía es preciso disponer de modelos animales fiables y seguros. En nuestro caso disponemos de dos tipos principales de modelos: los ratones transgénicos y los ratones trisómicos. En los transgénicos el ratón posee más de dos copias de un determinado gen, que opera por lo demás en un ambiente normal de disomía. En los trisómicos el ratón posee un tercer cromosoma o parte sustancial de él, un cromosoma que contiene regiones sinténicas de genes ortólogos con los del cromosoma 21 humano. Destaca particularmente el cromosoma 16 del ratón. Ambos modelos difieren sustancialmente pero, correctamente combinados, nos ofrecen una información complementaria de extraordinario valor para conocer muchos de los problemas que aparecen en el síndrome de Down. Voy a ponerles un ejemplo particularmente significativo.
 Diversos estudios mostraron que el gen DYRK1A participa en el crecimiento cerebral, tanto en ratones como en humanos. En ratones transgénicos, su sobreexpresión inhibe la proliferación neuronal en la corteza cerebral, incluido el hipocampo, e induce la diferenciación prematura de las células progenitoras neuronales. En estos ratones transgénicos, se aprecia una alteración en ciertas pruebas que analizan el aprendizaje y la memoria. Por otra parte, numerosos trabajos han mostrado también que diversos tipos de ratones trisómicos (todos los cuales presentan tres copias de Dyrk1A) muestran profundas alteraciones del aprendizaje y de la memoria. ¿En qué grado la sobreexpresión de este gen concreto puede contribuir a los problemas cognitivos del ratón trisómico? Sustraigamos al trisómico una copia de sólo el gen Dyrk1A, y veamos si mejora o no su capacidad cognitiva.
Diversos estudios mostraron que el gen DYRK1A participa en el crecimiento cerebral, tanto en ratones como en humanos. En ratones transgénicos, su sobreexpresión inhibe la proliferación neuronal en la corteza cerebral, incluido el hipocampo, e induce la diferenciación prematura de las células progenitoras neuronales. En estos ratones transgénicos, se aprecia una alteración en ciertas pruebas que analizan el aprendizaje y la memoria. Por otra parte, numerosos trabajos han mostrado también que diversos tipos de ratones trisómicos (todos los cuales presentan tres copias de Dyrk1A) muestran profundas alteraciones del aprendizaje y de la memoria. ¿En qué grado la sobreexpresión de este gen concreto puede contribuir a los problemas cognitivos del ratón trisómico? Sustraigamos al trisómico una copia de sólo el gen Dyrk1A, y veamos si mejora o no su capacidad cognitiva.
Para ello cruzamos ratones hembra trisómicas (Ts65Dn) con machos heterozigotos con una sola copia de Dyrk1A. Se obtuvieron tres clases de ratones: trisómicos con tres copias del segmento cromosómico en el que se encuentra Dyrk1A junto con otros genes (TS +/+/+), trisómicos para todos los genes de ese segmento salvo el Dyrk1A (TS +/+/-) y euploides con una dosis normal de Dyrk1A (CO +/+). Vean los resultados en la figura 1 que muestra la expresión de proteína, el nivel de aprendizaje en el test de Morris, y el grado de potenciación a largo plazo en circuitos del hipocampo.
Figura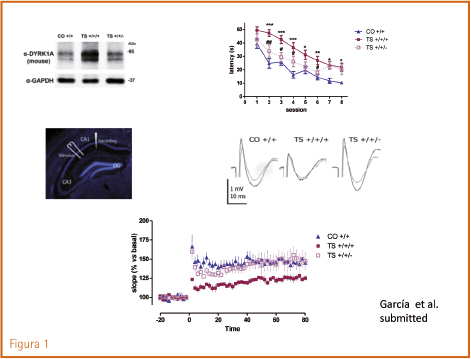 1. Expresión de proteína Dyrk1A, nivel de aprendizaje (test de Morris) y grado de potenciación a largo plazo en los tres grupos de ratones (control +/+, trisómicos +/+/+, y trisómicos a los que se les ha suprimido una copia de Dyrk1A +/+/-). (García et al., 2014).
1. Expresión de proteína Dyrk1A, nivel de aprendizaje (test de Morris) y grado de potenciación a largo plazo en los tres grupos de ratones (control +/+, trisómicos +/+/+, y trisómicos a los que se les ha suprimido una copia de Dyrk1A +/+/-). (García et al., 2014).
En resumen, nos indica que Dyrk1A: a) interviene en importantes funciones cerebrales que tienen que ver con la cognición, b) su sobreexpresión altera dichas funciones, y c) puede asumir una importante responsabilidad en los mecanismos funcionales del síndrome de Down.
Consecuencia: si consiguiéramos bloquear parte de la actividad DYRK1A en una persona con síndrome de Down, quizá pudiéramos mejorar su funcionamiento cognitivo. Esto es lo que se ha ensayado en España con la epigalocatequina, un alcaloide del té verde que inhibe la actividad cinásica de la proteína DyrkA, con ciertos resultados positivos.
Vean, por tanto, cómo la disponibilidad de modelos animales de síndromes que cursan con discapacidad intelectual nos van a permitir indagar y profundizar no sólo en los mecanismos que llevan a la discapacidad sino también a mejorarla. Sin duda, es uno de los grandes retos de la neurociencia en el siglo XXI.
Acción sobre la neuroquímica
Sin necesidad de llegar a manipular los genes y sus mecanismos de acción génica, ¿no podríamos también descubrir consecuencias fenotípicas de la trisomía, como por ejemplo las neuroquímicas, presentes en el cerebro del modelo animal, sobre las cuales pudiéramos actuar?
Una de estas consecuencias fenotípicas fue la constatación de que en determinadas zonas del cerebro trisómico, concretamente en el hipocampo, existe un predominio de la actividad inhibidora GABA, y se postuló que ello contribuiría a los problemas de aprendizaje y memoria observados en dicho animal y, por referencia, en el síndrome de Down. ¿Cabría frenar la acción inhibidora del GABA? Los antagonistas GABA-A de acción generalizada mejoran la cognición en este modelo, pero su capacidad de antagonizar todos los subtipos de receptores GABA-A tiene el riesgo de producir convulsiones, ansiedad, insomnio, etc. ¿Sería posible disponer de un producto antagonista con afinidad específica por un subtipo de receptor GABA-A, el llamado α5, que se encuentra prioritariamente en el hipocampo y no se distribuye tan extensamente por todo el cerebro? La respuesta fue, sí. El Ro-4938581 es selectivo para 5 en donde actúa como agonista inverso y por tanto contrarresta la acción del GABA. Conseguido el producto, ¿qué acciones ejerció en el ratón trisómico? Se exponen en la figura 2. La acción del agonista inverso mejoró de forma importante la habilidad cognitiva, revirtió la reducción de la LTP y mejoró la neurogénesis cerebral.
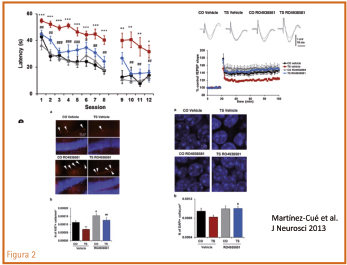 Figura 2. El agonista inverso 5, Ro-4938581, mejoró la cognición en ratones Ts65Dn (test de Morris), el grado de potenciación post-tetánica (LTP) en el hipocampo, y mejoró la neurogénesis en hipocampo. (Martínez-Cué et al., 2013).
Figura 2. El agonista inverso 5, Ro-4938581, mejoró la cognición en ratones Ts65Dn (test de Morris), el grado de potenciación post-tetánica (LTP) en el hipocampo, y mejoró la neurogénesis en hipocampo. (Martínez-Cué et al., 2013).
Consecuencia. Controlar el exceso de actividad GABA de forma selectiva en determinadas regiones del cerebro puede resultar beneficioso para las personas con síndrome de Down. Los laboratorios Roche, creadores de estas moléculas, han iniciado un ensayo clínico en el que están participando varios países para comprobar sus efectos en personas con síndrome de Down.
Organización funcional
Un último apunte. Numerosos estudios han demostrado que en el cerebro del síndrome de Down existe una clara reducción tanto en el número de unidades neuronales como en el de sus prolongaciones dendríticas y neuríticas, especialmente en el cerebelo, corteza prefrontal, hipocampo y lóbulo temporal. Es consecuencia de alteraciones en los sistemas de proliferación y diferenciación, ya perceptibles en el periodo embrionario, que se mantienen a lo largo de la vida. Junto a estas evidencias estructurales, se han demostrado también problemas en los mecanismos de señalización propios de la transmisión neuroquímica. Este déficit del aparato por donde debe fluir la información, que se inicia en las etapas tempranas del desarrollo cerebral, necesariamente ha de repercutir en la formación, instauración y mantenimiento de las redes y circuitos funcionales que subyacen y operan en las diversas funciones del cerebro. Las técnicas de neuroimagen funcional nos ayudarán a esclarecer esta realidad. Vean un ejemplo obtenido mediante resonancia magnética funcional por el grupo de Schapiro, estudiando el procesamiento del lenguaje en su vertiente auditiva: (figuras 3 y 4).
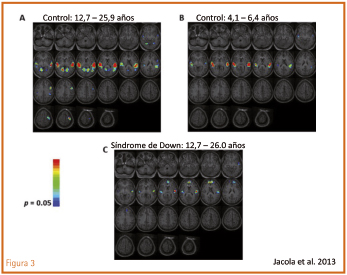 Figura 3. Imágenes de resonancia magnética en cerebros: A) grupo con desarrollo ordinario de la misma edad cronológica que la del síndrome de Down; B) grupo con desarrollo ordinario de la misma edad mental que la del síndrome de Down; C) grupo con síndrome de Down. Las imágenes se presentan de acuerdo con la convención radiológica, de modo que el lado izquierdo de la imagen corresponde al hemisferio derecho del cerebro. (Jacola et al., 2013).
Figura 3. Imágenes de resonancia magnética en cerebros: A) grupo con desarrollo ordinario de la misma edad cronológica que la del síndrome de Down; B) grupo con desarrollo ordinario de la misma edad mental que la del síndrome de Down; C) grupo con síndrome de Down. Las imágenes se presentan de acuerdo con la convención radiológica, de modo que el lado izquierdo de la imagen corresponde al hemisferio derecho del cerebro. (Jacola et al., 2013).
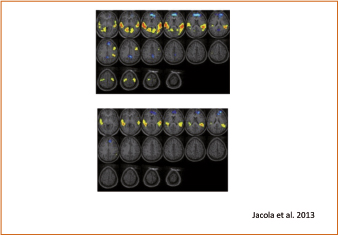 Figura 4. Análisis inter-grupos entre el grupo con síndrome de Down y los grupos con desarrollo ordinario de la misma edad cronológica (a) y de la misma edad mental (b). Las regiones en amarillo-naranja corresponden a áreas en las que el grupo con desarrollo ordinario fue activado más significativamente que el grupo con síndrome de Down. Regiones en azul ligero a oscuro corresponden a áreas en las que hubo activación significativamente mayor en el grupo con síndrome de Down que en los grupos con desarrollo ordinario. Las imágenes se presentan de acuerdo con la convención radiológica, de modo que el lado izquierdo de la imagen corresponde al hemisferio derecho del cerebro. (Jacola et al., 2013).
Figura 4. Análisis inter-grupos entre el grupo con síndrome de Down y los grupos con desarrollo ordinario de la misma edad cronológica (a) y de la misma edad mental (b). Las regiones en amarillo-naranja corresponden a áreas en las que el grupo con desarrollo ordinario fue activado más significativamente que el grupo con síndrome de Down. Regiones en azul ligero a oscuro corresponden a áreas en las que hubo activación significativamente mayor en el grupo con síndrome de Down que en los grupos con desarrollo ordinario. Las imágenes se presentan de acuerdo con la convención radiológica, de modo que el lado izquierdo de la imagen corresponde al hemisferio derecho del cerebro. (Jacola et al., 2013).
Comentarios
Los resultados demuestran que, en el procesamiento del lenguaje oído o escuchado, existe un patrón diferente de activación cerebral en los jóvenes adultos con síndrome Down al compararlos con grupos con desarrollo ordinario de su misma edad cronológica o mental. En los grupos con desarrollo ordinario, la activación neural siguió un patrón bien definido y clásico, caracterizado por la fuerte activación de la corteza auditiva en ambos hemisferios, que refleja el procesamiento inicial de la información auditiva. Es lógica la diferencia en la intensidad de activación, que fue mayor en el grupo adulto (fig. 3) que en el grupo infantil (fig. 4). Se aprecia también un proceso de lateralización de la información hacia el hemisferio izquierdo en las regiones frontal y parietal, conforme la edad avanza y se desarrolla el lóbulo frontal. Estas regiones frontales son las que mantienen la información online (memoria operativa verbal) y el procesamiento sintáctico.
Los jóvenes con síndrome de Down mostraron un patrón de activación cualitativa y cuantitativamente diferente. No se vio activación en las regiones del lóbulo frontal izquierdo durante el procesamiento del relato; es decir, se vio lo mismo que ocurrió con el grupo de menor edad con desarrollo ordinario, lo que sugiere que la maduración frontal en el síndrome de Down puede estar retrasada; o bien que, como indican los estudios estructurales de esa región en el síndrome de Down, hay una alteración en la región frontal y en su función. Puesto que son regiones relacionadas con la memoria operativa verbal y el procesamiento sintáctico, eso explicaría las alteraciones que las personas con síndrome de Down padecen en tales funciones, que no les permiten procesar con la debida rapidez y fluidez la información verbal que reciben, interfiriendo así el complejo desarrollo y progreso del habla. Lo mismo cabe decir al analizar la pobre activación observada en este grupo, en comparación con la de sus pares por edad cronológica o mental, en las regiones de la corteza temporal y parietal de ambos hemisferios, áreas que son particularmente activas en las tareas que implican el procesamiento del lenguaje. De nuevo, la causa puede residir en la menor riqueza de conexiones y conformación de redes neuronales dentro de estas áreas en el cerebro del síndrome de Down.
En cambio, el grupo síndrome de Down mostró una importante activación en las regiones de la línea media del lóbulo frontal y en la corteza cingulada, algo que no ocurrió en los dos grupos con desarrollo ordinario. Una posible interpretación podría ser la diferencia en el modo de ejecutar una tarea. Si la tarea es compleja, los grupos con menor capacidad de ejecución pueden desarrollar un aumento compensador en la activación de regiones cerebrales frontales relacionadas con funciones ejecutivas. No parece ser el caso, dado que la tarea era lo suficientemente sencilla como para poder ser bien ejecutada por niños pequeños. Cabe otra interpretación: que se trate de una actividad añadida, puesta en marcha por la estimulación auditiva, que resulte disfuncional y perturbe el procesamiento de la información auditiva en las áreas específicamente responsables de dicha tarea.
En  definitiva, lo que se observa en jóvenes adultos con síndrome de Down es la presencia de unos patrones atípicos de activación y funcionamiento en regiones cerebrales relacionadas con el lenguaje: algunas de estas regiones eran claramente menos activas y, en cambio, aparecieron otras alternativas. Es decir, se aprecia una marcada diferencia en la organización de las redes neuronales responsables de recibir e interpretar la información auditiva de un relato. Nótese, sin embargo, que al final la información llega y se hace consciente; pero lo consigue por procedimientos y mecanismos diferentes, aparentemente más sinuosos y débiles. El estudio es realizado en jóvenes adultos de edades entre 12 y 26 años. Parece lógico pensar que esta disfunción organizativa, debida a la menor riqueza de redes neuronales en los sitios críticos de procesamiento del lenguaje, se encuentra ya presente desde la infancia y la niñez, justo en las etapas que son vitales para iniciar dicho procesamiento. Tal puede ser una de las causas por las que se aprecian marcadas dificultades en el aprendizaje y posterior utilización del lenguaje y del habla en las personas con síndrome de Down.
definitiva, lo que se observa en jóvenes adultos con síndrome de Down es la presencia de unos patrones atípicos de activación y funcionamiento en regiones cerebrales relacionadas con el lenguaje: algunas de estas regiones eran claramente menos activas y, en cambio, aparecieron otras alternativas. Es decir, se aprecia una marcada diferencia en la organización de las redes neuronales responsables de recibir e interpretar la información auditiva de un relato. Nótese, sin embargo, que al final la información llega y se hace consciente; pero lo consigue por procedimientos y mecanismos diferentes, aparentemente más sinuosos y débiles. El estudio es realizado en jóvenes adultos de edades entre 12 y 26 años. Parece lógico pensar que esta disfunción organizativa, debida a la menor riqueza de redes neuronales en los sitios críticos de procesamiento del lenguaje, se encuentra ya presente desde la infancia y la niñez, justo en las etapas que son vitales para iniciar dicho procesamiento. Tal puede ser una de las causas por las que se aprecian marcadas dificultades en el aprendizaje y posterior utilización del lenguaje y del habla en las personas con síndrome de Down.
Me encantaría mostrar más estudios. Por ejemplo, cómo se está analizando en modelos murinos de autismo el modo en que los ratones se comunican, uno de los grandes problemas de esta forma de discapacidad. Y siguiendo el mismo esquema, la comunicación en ratones modelo de síndrome de Down, en donde la comunicación es también uno de sus principales problemas. Pero el tiempo no da más de sí.
En conclusión
He tratado de mostrar simples esbozos de cómo el estudio cerebral de la discapacidad intelectual enriquece nuestro conocimiento sobre el funcionamiento de nuestro cerebro. Pero, al mismo tiempo, nos invitan a pensar en lo que la neurociencia puede y debe aportar al mundo de la discapacidad intelectual. Es su gran reto. No sólo se trata de conocer mejor la entraña del cerebro y de su funcionamiento sino de, a partir de ahí, encontrar medios que mejoren la discapacidad. En definitiva: conocer más para servir mejor.